核酸・蛋白質科学研究室(B)
(新井)
Laboratory of Molecular and Cellular Biochemistry (B) [LMCB-B]

研究テーマ

「真核生物の遺伝子組換えの分子機構の研究」
ゲノム編集ということを聞いたことがあると思います。CRISPR-CAS9という酵素(タンパク質とRNAの複合体)を使うと簡単に生物のゲノムDNAを書き換えられるというイメージでしょうか。では、CRISPR-CAS9って何をしているのか知っていますか?CRISPR-CAS9が行っているのは、DNAの狙った塩基配列を切断するということです。ただそれだけ。(とはいえ、確かに狙ったDNA塩基配列を切断できるのは画期的ではあります)それでは、なぜゲノム編集ができるのでしょう?CRISPR-CAS9によりDNAが切断された後は、生物が持つ機能によって切断箇所が修復され、多くの場合は元に戻ります。しかしその一部でDNAの組換えが起こって変化が生じ、人間はその中から都合の良いものを選別しているだけです。もっと効率よくできればいいのですが、、、。それにはDNAが切断された後に、どのように修復されたり組換えが起こっているのかについて、メカニズムを解明して、人間がコントロールできるようになる必要があります。
生物が自ら制御して行うDNAの組換え(再編成)には以下のようなものがあり、必ずDNAの切断(二本鎖DNAの切断)が伴います。
減数分裂期組換え [DNAの再編成]
我々の遺伝子 DNAは正確に複製され、そして子孫へと受け継がれています。しかし、生殖細胞 を形成する過程でDNAが再編成されています(減数分裂期組換え)。この減数分裂期組換えでは、生物が自らDNAを切断することから始まり、その切断を生物自身が修復しながら組換えを行っています。この時のDNAの再編成には「相同組換え」が働きます。
[DNAの修復]
普段の生活の中では、宇宙放射線、強い紫外線、化学物質等によってDNAの切断が起こっています。
その切断された DNAを生物は修復して元の状態に戻しています。この時に、DNAが組換わってしまうことがあります。この時の二本鎖DNAの切断された時、「相同組換え」と「非相同末端結合」が働きます。
DNA切断から始まる「DNAの再編成」と「DNAの修復」には生物共通のメカニズムがあります。それが「相同組換え」です。相同組換えは、 DNA修復という「正確性」とDNA再編成という「多様性」の両方に関わっています。一方、DNA切断の修復には「非相同末端結合(NHEJ)」というもう一つの機能があります。非相同末端結合はDNA切断の修復に働きますが、減数分裂期組換えには働きません。
当研究室では、生体内で起こっているDNA 組換えのメカニズムを解明するために、関連する遺伝子からタンパク質を人工的に作らせて精製し、それらのタンパク質を試験管内で再構成する生化学的手法を中心に、 遺伝子の機能を改変した細胞を用いた分子遺伝学的手法、DNAとタンパク質の複合体を直接観察する走査型プローブ顕微鏡 SPMによるナノテクノロジーを駆使しながら研究を進めています。


相同組換えの分子メカニズム
相同組換えのDNAレベルでの分子メカニズムは,(1) DNAの切断,(2) 相同なDNA鎖の検索対合,(3) 再結合の過程に分けることができます。その中で(2) の相同な(塩基配列の似ている)DNA鎖を探し出し対合する過程を「相同対合」(homologous pairing) と言います。
私たちは、「細胞内の長大なゲノムDNAから如何に塩基配列の似ている部位を探し出すのか?」という点に着目しています。まず、相同組換えに関与している遺伝子から人工的にタンパク質を作り、それを精製単離します。続いて、それらのタンパク質と2種類のDNAを試験管の中で混ぜ合わせて生体内での相同対合反応を再現させます。このような生化学的方法を中心に用いて、相同組換えに関与しているるタンパク質の機能を明らかにすることにより、相同組換えの分子メカニズムの解明を目指しています。

相同組換えを試験管の中で再現
相同対合を試験管の中で解析するために、一本鎖DNA断片(ssf)と閉環状二本鎖DNA(form I)を基質とした反応系を使います。その反応系に組換えタンパク質を加えて相同対合が形成されると "D-loop" と呼ばれる組換え中間体が形成されます。
真核生物の相同組換えの研究には出芽酵母は最適です。
出芽酵母 (Saccharomyces cerevisiae)
・真核生物である。
・ヒトと相同遺伝子がある。
・高等生物のモデル系となり得る。
・相同組換えの効率が高い。
・ゲノムDNAの情報が豊富。
・ゲノムDNAの改変が容易。
・培養も容易(お金がかからない)。
出芽酵母の相同組換えに中心的な役割を担っているタンパク質がRad51とRad52です。

組換え中間体 [D-loop] を捕える
相同対合によって形成された D-loop は電気泳動により分析されます。放射性物質または蛍光物質で標識した直鎖状一本鎖DNA断片(ssf)と閉環状二本鎖DNA(form I)を用いて反応を行い、アガロースゲル電気泳動でDNA を分離します。ssfは速く移動し、form Iはゆっくり移動します。D-loopはssfとform Iとの複合体ですから、分析機器イメージアナライザーTyphoonで放射性物質や蛍光物質を解析すると、D-loopはform Iの位置に現れるssfのシグナルとして検出されます。。
私たちは、出芽酵母のRad51タンパク質とRad52タンパク質の両方がある時にD-loopが効率よく形成されることを発見しました。
大腸菌のRecAタンパク質は単独でD-loop形成を触媒します。Rad51とRecA は働きがよく似た相同遺伝子です。
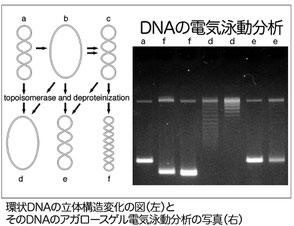
DNAの形状変化を解析
組換えタンパク質がDNAに結合すると、DNAに様々な構造変化を与えます。この変化をアガロースゲル電気泳動で分析します。上の結果は、Rad51が二本鎖 DNA に結合するとDNAの二重らせんを緩める(e, f)ということを環状DNAの超らせんの変化により解析した一例です。

Rad51は相同性検索と対合を行う
Rad51はDNA上に連続して結合します。これをフィラメント構造(nucleoprotein filament) といいます。Rad51は一本鎖DNAと二本鎖 DNA の相同性(塩基配列の似ている領域)を探し出して、一本鎖DNAを二本鎖 DNA の間に侵入させて、相補鎖と水素結合を形成させるための中心的役割をしています。その時にD-loopが形成されます。上の図は、Rad51とRad51が結合した立体構造で、関連するアミノ酸(赤、青)が示されています。黄緑はDNA結合に関連するアミノ酸です。図中の右下の写真は二本鎖 DNA 上に形成されたRad51 フィラメントを走査型プローブ顕微鏡で観察したものです。

Rad52は酵母の相同組換えに必須
Rad52は出芽酵母の相同組換えに最も重要なタンパク質です。Rad52がないと相同組換えが全く起こらなくなります。
多くの働きが知られています。
- 一本鎖DNAに結合しているRPA(一本鎖DNA結合タンパク)をRad51と交換する働き。
- Rad51と複合体を形成して、二本鎖 DNA を捕まえてRad51に渡す働き。
- Rad51のD-loop形成を促進する働き。
この他に、Rad52は単独でも働いていますし、まだまだわかっていない働きも多くあると考えられています。



研究室の実験機器、装置
左:AKTA、カラムクロマトグラフィーによりタンパク質を精製する装置(低温室4˚Cに設置)
中央:ゲル濾過カラム(分子ふるい)Shaphacryl S300HR、赤(上)チトクロームC、青(カラム下方(薄い))ブルーデキストラン。
右:GelDoc、DNA・タンパク質の電気泳動などの画像解析装置
[Enjoy Science !]