タンパク質工学的手法による P450 イソフラボノイド骨格合成酵素の
反応機構と分子進化の研究
澤田 有司
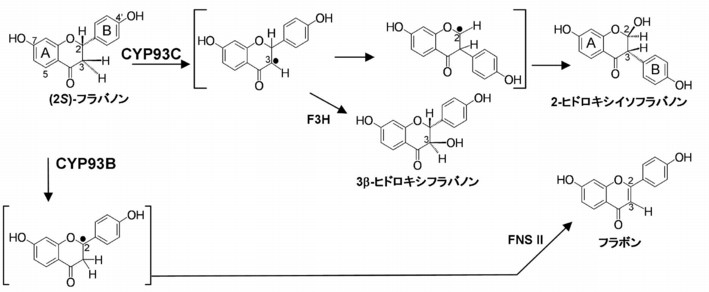
マメ科特有のP450分子種CYP93C は,基質 (2S)-フラバノンからアリール基の1,2-転位を伴う2位へのヒドロキシル化反応によってイソフラボノイド生合成の初発物質2-ヒドロキシイソフラバノンを生成する(Fig. 1).イソフラボノイドは抗菌活性や動物へのエストロゲン作用などを示す防御物質として働く一方,一部のマメ科では根粒形成シグナルとしても働く.このような特殊な P450 反応が,マメ科植物の生態生理にとってきわめて重要なイソフラボノイド生合成の鍵段階となっていることから,この反応機構を解明し,触媒作用に必要なタンパク質構造とその分子進化の過程を明らかにするために以下の研究を行った.
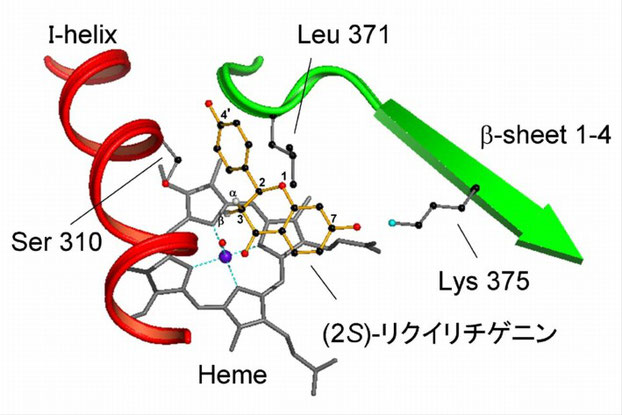
マメ科カンゾウの CYP93C2 の推定タンパク質構造を構築し,基質と相互作用しうる部位を調べた (Fig. 2).次に,推定された鍵アミノ酸残基候補を部位特異的変異導入で他のアミノ酸残基に置き換え,酵母異種発現系で機能を調べた.その結果,タンパク質構造の維持に関係する部位,P450 反応サイクルで酸素分子の活性化に必須な部位,反応特異性にかかわる部位などが見出された.特に,Ser 310はアリール基が転位する先に位置しこの部位の空間的余裕が転位反応に重要であること,Lys 375 は基質の7位ヒドロキシル基と相互作用しうる位置に存在し転位反応に必須であること,基質の1位酸素近傍に存在する Leu 371 はタンパク質構造の維持と転位反応に適した位置への基質の結合に重要であることがわかった.これらの部位の変異酵素は著しく反応特異性が変化し,Lys 375 を Thr に置き換えた K375T は転位反応が全く見られず,フラバノン 3-ヒドロキシル化酵素 (F3H) 活性のみを示した.さらに,3箇所変異を組み合わせた S310T-L371V-K375T による反応では,基質に二重結合が導入されたフラボンのみが生成した.興味深いことに,これらの活性を持つ変異酵素は野生型より熱安定性が増加していた.
CYP93 ファミリー間の系統樹では CYP93C と非マメ科植物に広く分布するフラボン合成酵素 (CYP93B) が同じグループに分類される.CYP93C への複数の変異導入によってフラボン合成活性が新生したことより,次のような分子進化が推定された.先祖型のフラボン合成酵素活性を持つ P450 より現存の CYP93B が生じる一方,遺伝子重複と上記鍵アミノ酸残基を含む変異の蓄積により,基質の結合様式が微妙に変化し,P450 型の F3H を経て新たなアリール基転位という反応特異性が獲得された.また,この過程ではタンパク質構造の安定性を犠牲にして新規の機能が獲得された.そして,生産物イソフラボノイドの生体防御や共生など生態系での有利な生理活性により,マメ科に機能が固定されたと思われる.
今後,CYP93Cのより精度の高い立体構造情報に基づいた反応機構の解明が望まれる.また,他の植物にも存在する機能未同定のCYP93の酵素機能と生理機能が明らかになれば,CYP93の分子進化の全体像が示されるものと期待している.